Laboratory of group mind
This chapter serves as a short and friendly introduction to the formalism used throughout the thesis. As discussed in the introduction, a persistent tension runs through the social sciences: methodological individualists seek to explain social behavior by reducing it to individual preferences, beliefs, or values, while group realists argue that social groups possess irreducible properties that shape outcomes in their own right.
This tension reappears in modeling, though often in subtler ways. Schumpeter's focus on individual decision-making helped lay the groundwork for rational choice theory and game theory §1.1. Meanwhile, social scientists inspired by Simmel's idea of duality used bipartite graphs to represent individuals and their affiliations. While such models acknowledge social context, they often treat groups as metadata on individuals rather than as entities with dynamics of their own. Hence, these models fail to capture the multiscale feedback between individuals and institutions, which could be thought of as a stronger reading of Simmel's duality.
One domain where this modeling tension is especially clear is the problem of cooperation (§1.1.1). If evolution favors selfish individuals---whether rational agents or self-replicating genes---how does large-scale cooperation emerge, especially among unrelated individuals? A common answer is positive assortment: cooperators tend to interact more often with one another than with defectors. This thesis can be read as a modeling ontribution to that broader puzzle.
We close the chapter by introducing the master equation formalism used in Chapter GroupSkills, along with its group-based extension developed in Chapter Coevo.
Abstract
We combine two families of models to explore the complexity of groups and institutions; contagion models and evolutionary game theory models. Both have recently been extended using hypergraph and approximate master equation formalism to represent group dynamics. We first introduce mean-field approaches, how they are used in evolutionary game theory, before moving to (approximate) master equations.
Mean-field theory
Many classical models of population biology, epidemiology, and the
social sciences rely on mean-field theory, which assumes that each part
of the system interacts only with the average or expected state of the
others---thereby ignoring structural and dynamical correlations. In this
approximation, local fluctuations and heterogeneity are smoothed out,
allowing for simpler, often analytically tractable models. A well-known
example is the Lotka-Volterra model, where predator-prey interactions
are modeled as random encounters between a predator population,
where
The problem of cooperation
The prisoner's dilemma (PD) epitomizes how cooperation was problematized in the social sciences. In classical games, the story is about a pair of individuals who must choose their next move, weighing their choice with an underlying utility function that specifies their preferences over every action profile. Players are assumed to be consistent in their utility functions and knowledge of their opponent's strategy options. Beyond the story that we will not recite here, PD is about the following (generic) payoff matrix
where
The Nash equilibrium is a simple mathematical result, but it has often been interpreted as proof that self-interested individuals must be tamed---since no one will unilaterally deviate from mutual defection without external incentives [@smaldino_modeling_2023]. By "tamed", we mean that later models showed how cooperation can emerge when agents face punishment threats, such as state intervention. Although we now know there are many ways out of the Prisoner's Dilemma, this line of reasoning influenced scholars like Garrett Hardin in formulating the tragedy of the commons: the idea that rational individuals will overuse and deplete shared resources, such as fisheries or grazing land [@hardin_tragedy_1968]. One could say that simple models of rationality--by assuming selfishness--helped create their own tragedy: shaping institutions that expect defection, discourage trust, and crowd out local cooperation.
The evolution of cooperation
Whereas classical games frame the problem of cooperation in terms of strategies, rationality, and mind reading, evolutionary game theory examines populations where behavioral strategies are inherited, and individual fitness is determined by how common these strategies are within the population. That is, evolutionary game theory is concerned with situations where the consequence of a variable characteristic depends on its frequency in the population, or frequency dependence. In this thesis, we will be particularly interested in how frequency dependence in structured population importantly define the scope of group interactions. Stated bluntly, group interactions can be thought as the fitness of certain traits, not people, based on the joint influence norms and individual psychology in a group-structured population.
Evolutionary game theory is rooted in genetics, with alleles (or genotypes) co-varying with heritable behavioral programs, or strategies [@hamilton_genetical_1964]. A key simplifying assumption is that single alleles code for behaviors. We will later discuss cultural variants, typically transmitted through social transmission. Social learning can occur in multiple directions---vertically (from parents to offspring), horizontally (among peers), or diagonally (from relatives such as uncles). Humans are unique in that individuals also adopt behaviors by emulating those who are perceived as more successful or prestigious, referred to as success- and prestige-biased transmission. But for now, it is useful to understand where the notion of fitness comes from before applying it to cultural systems. In Chapter 4, we explore the consequences of groups copying the institutions of more successful groups, based on perceived fitness within a contagion context
The replicator dynamics is a key result in evolutionary game theory that will allow us to connect literature from cultural evolution with that of public good games (PGGs) on networks
where
To illustrate the usefulness of the replication dynamics, consider the following Hawk-Dove (or Snowdrift) game, where at each round a pair of individuals fights over a non-divisible resource. In this case, the Hawk and Dove strategies can be summarized with the following payoff matrix
where }, while the expected payoff for a Dove is $W(D)=p(0) + (1-p)V/2}. A central question in evolutionary game theory
is whether these strategies can coexist in the long run as evolutionary
stable states (ESS). The payoffs are linked to evolutionary fitness, as
higher payoffs result in passing down the strategies to subsequent
generations. To answer this question for a particular strategy, say
Hawk, we solve for all values of
A well-known result is that
Positive assortment and complex networks {#sub.assort}
Both classical and evolutionary game theory suggest that randomness tends to favor the greedy. In a sea of hawks, doves are at a disadvantage. But if doves preferentially seek out other doves, they gain a foothold---an effect known as positive assortment. Nowak & May (1992) demonstrated a version of this in grid-based Prisoner's Dilemma (PD) games, showing that cooperator clusters can emerge to promote cooperation [@nowak_evolutionary_1992]. Yet, Hauert and Doebeli (2004) later observed that spatial structure does not always enhance cooperation; in a grid with uniform neighborhoods, a higher cost-to-benefit ratio is required than in an unstructured population [@hauert_spatial_2004]. These results motivated researchers to investigate more realistic forms of social structures, including heterogeneity in social contacts, to understand their impact on cooperation [@santos_scale-free_2005; @ohtsuki_simple_2006].
In many real-world social networks, a small number of individuals have disproportionately many connections, while most have relatively few---producing a heavy-tailed degree distribution [@simon_class_1955; @barabasi_emergence_1999]. To capture such heterogeneity, one can group nodes into compartments based on shared attributes (e.g., degree, but also social status, age group, or even evolutionary strategy). Rather than tracking a single, fixed realization of the network, an annealed approximation is often used, wherein links are constantly reshuffled at a rate much faster than the dynamics of interest (e.g., strategy updates or contagion). This constant "reshuffling" of connections means we can effectively work with an ensemble of possible networks that obey certain constraints, thus reflecting these constraints on average, without being locked into any one realization.
Under the annealed approximation, a random contact of a node in
compartment
where
A well-known mechanism for generating skewed or power-law degree distributions is the preferential attachment growth process [@simon_class_1955; @barabasi_emergence_1999]. In this model, new nodes are more likely to connect to already well-connected nodes, a "rich-get-richer" phenomenon. One common form of the resulting degree distribution is:
where
since for large
Limitations of the mean-field theory
All of the results so far rely on the mean-field assumption, which has its limitations. First, when one population's fequency is too high relative to the other, it can drive the other population to very low levels. However, because the mean-field approach averages over the entire population and assumes smooth, deterministic dynamics, both populations can approach zero asymptotically without ever exactly reaching zero. This occurs provided that the initial conditions and model parameters are such that neither population collapses to zero immediately. The smooth nature of these models does not account for individual-level fluctuations or other factors that could lead to sudden extinction.
Even though individuals interact on complex networks, modeling the dynamics often assumes random mixing, where the interaction network is reshuffled much faster than the unfolding of the dynamics. In this framework, interactions are random, but the units involved can have different characteristics that influence how frequently they interact with each other. Because these interactions are reassigned at each time step, the states of different units become independent over time, "washing out" any correlations between their behaviors. For example, even though Alice, Bob, and Trent might interact more often than others due to their age, they may also form a tightly-knit clique, influencing one another's states in nontrivial ways---an effect that the annealed approximation fails to capture.
To better account for such local dynamical correlations arising from the joint influence of individual node states and higher-order group interactions, we turn to master equations. These equations track the evolution of individual unit states while preserving information about the structure of interactions within the network, offering a more precise description of the system's dynamics than the mean-field approach, which only considers averaged behaviors. In particular, group-centered approximate master equations allow us to describe group-based processes more accurately by shifting focus from individual nodes to classes of groups, distinguished by their size and by the number of cooperators within them.
Modeling with the master equation
With master equations, we are concerned with the probability of the system being in a specific configuration at any given time. That is, rather than asking how the average number of cooperators or defectors changes over time, we care about the full range of possible interactions and how they evolve. For instance, having a configuration with five cooperators and one defector is not necessarily equivalent to its inverse. The exact arrangement matters: a lone defector among cooperators might behave differently than a lone cooperator among defectors. Master equations allow us to keep track of such distinctions. As such, they are exact, in contrast to the group-based master equations we introduce next.
We examine Weidlich and Haag's (1983) Lotka-Volterra master equation model, which extends the prey-predator dynamics to include a refuge habitat where prey are out of reach of predators (Fig. 1.4) [@haag_modelling_2017]. In this model, they show how the refuge habitat contributes to modeling the instability of the Lotka-Volterra cycles against perturbations, resulting in stable cycles instead of unstable closed curves. The modeling of different habitats, and how we can model the effect by taking into within--group composition, as well as the influence of groups interacting with each other, will be a recurrent theme in this thesis.
We represent the system as a time-dependent distribution over states
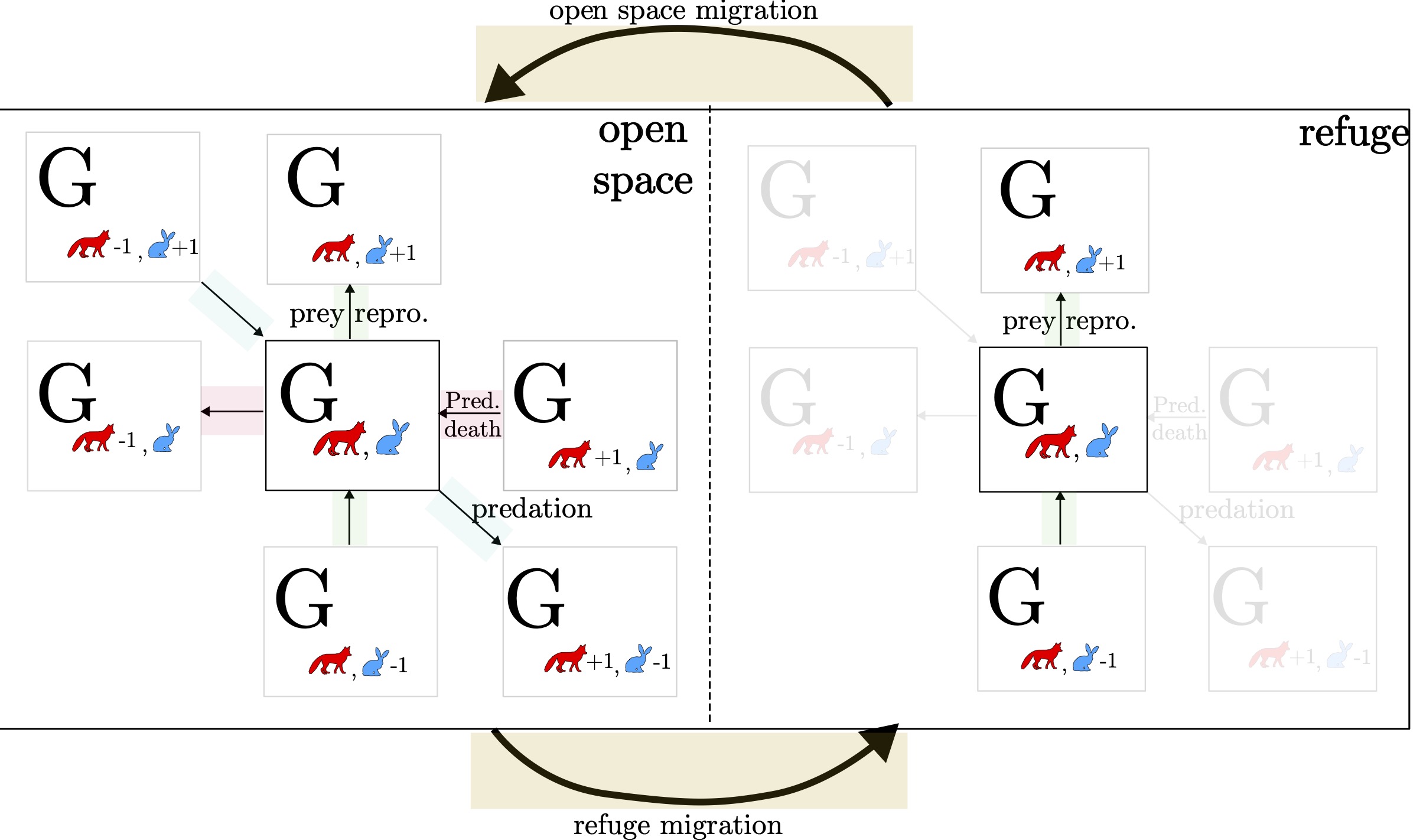
In Fig. 1.4, each compartment represents a state of the entire system. That is, each 'state' corresponds to different configurations, or ensembles, of the (multiverse) system, allowing us to model the probability distribution over all the possible configurations of the system over time. As before, the model retains the same basic information---predation, prey reproduction, and predator deaths---but now we add more details. Specifically, we introduce two distinct habitats in the system. These habitats may influence how the populations interact, which will become important as we model the system's dynamics in more depth. We provide the relevant bits from before but here written as master equations
But master equations are all about balancing our equations so that we
have normalization and conservation of our densities. That is, our
set of differential equations respects the
We do the same work for the other parts of the model in the appendix. The important thing is that in the end, we can combine the different parts of the model to get the following master equation:
In figure 1.5, we visualize the systems by either looking at the time dynamics (top rows) or examining the systems at equilibrium (bottom row). The upper left figure demonstrates what is meant by time evolution dynamics, where we can see how the states with 25 predators in the system are becoming increasingly more likely over time. In the upper right figure, we plot the expectation for the different populations, showing
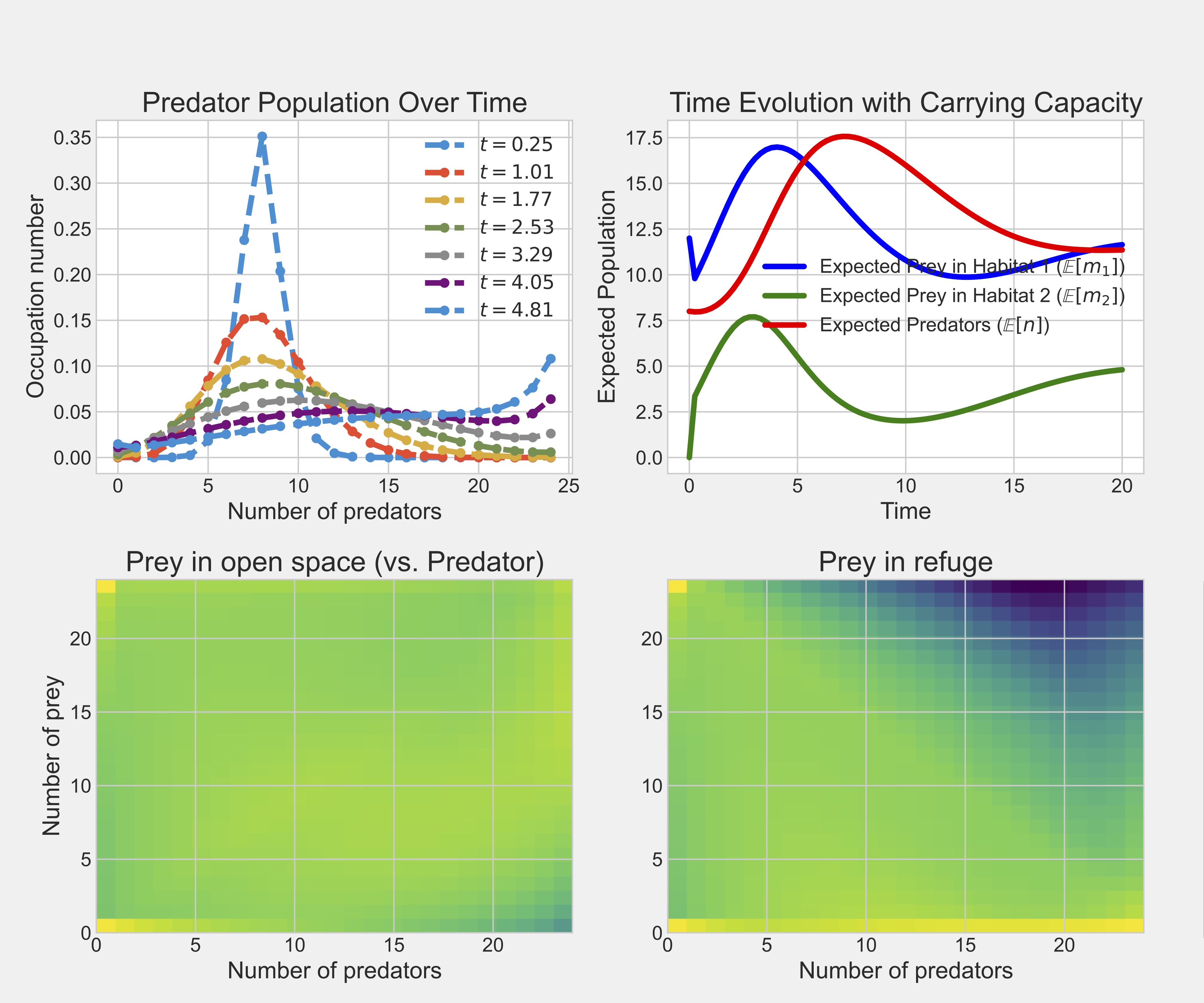
With the probability distributions in hand, we can calculate, for
instance, the expected population in the open space habitats by summing
over the entire probability distribution
Group-based master equations
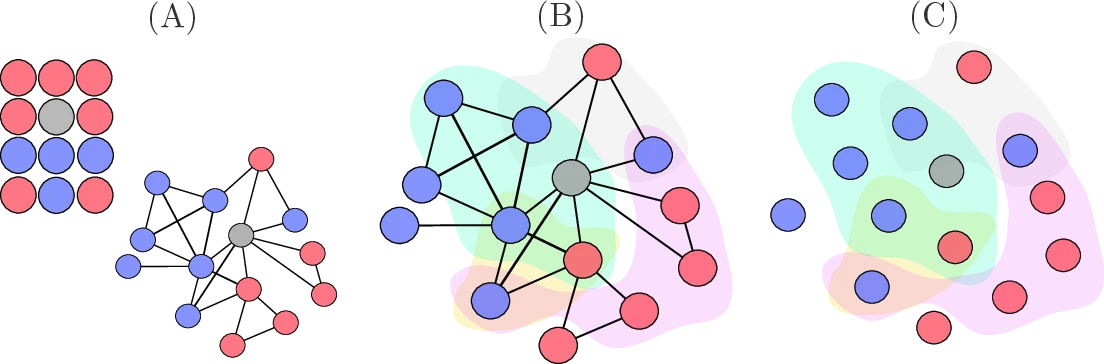
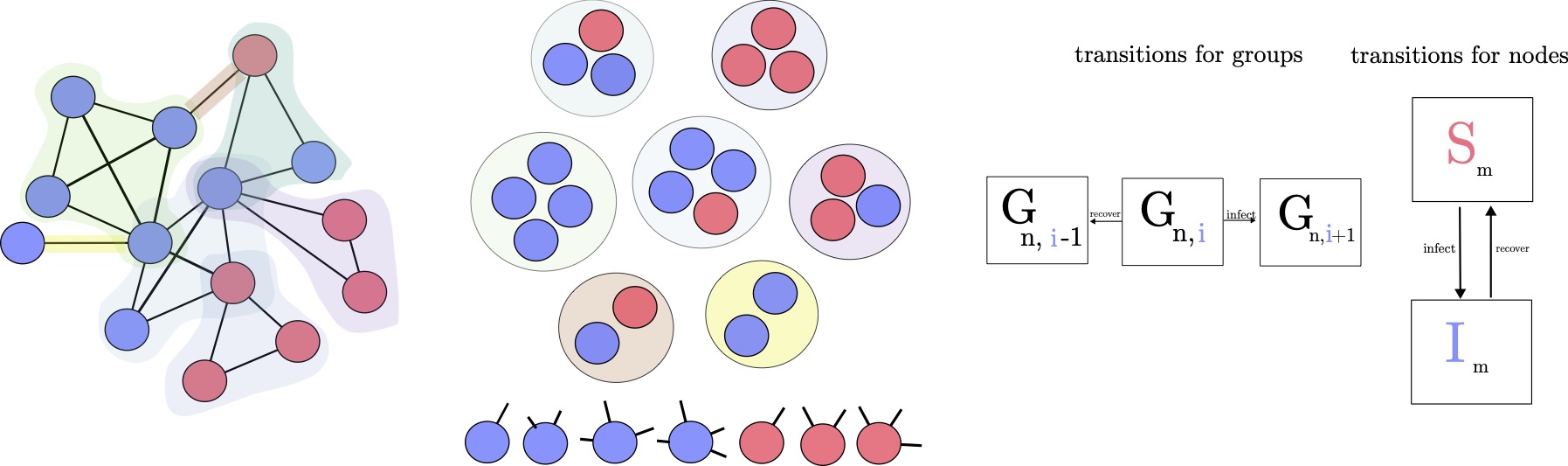
By coarse-graining our set of master equations around relevant subunits, we reduce the system's dimensionality while preserving its core dynamics. We distinguish between equations for individual members---capturing transitions between states within groups--and equations for the groups themselves--tracking how entire group configurations evolve over time. In Fig. 1.6, subunits are shown as circles, which may represent cliques or hyperedges. For instance, we can write 7 member equations to model the internal interactions among group members. These provide a more fine-grained description of the system, whereas group equations summarize how group-level states evolve. We refer to this formalism as group-based master equations (GMEs), in contrast to node-centered master equations where the approximation depends on the states of nodes and their neighbors [@marceau_adaptive_2010].
Building on previous work on approximate master equations [@hebert-dufresne_propagation_2010; @st-onge_master_2021; @st-onge_influential_2022], we extend beyond traditional two-species dynamics (e.g., prey-predator models) to group-based contagion, where individuals transition between Susceptible and Infected states (SIS dynamics). Unlike mean-field models that wash out correlations between individual states, GMEs preserve the exact composition of each group during contagion. This is crucial in settings like airborne virus spread within households or workplaces, where repeated exposure and shared environments lead to strong local reinforcement and non-linear transmission effects.
Coupling between groups arises naturally when individuals belong to multiple groups, creating pathways for contagion or influence to spread indirectly across the system. Originally introduced to model context-dependent contagion [@hebert-dufresne_propagation_2010], this approach has since been applied to settings such as policy co-evolution and collective behavior [@hebert-dufresne_source-sink_2022]. In our framework, such overlap induces dynamic coupling without requiring direct group-to-group interactions--capturing the essence of coupling from our typology in the Interface chapter
We highlight the different places where infection occurs in pink. First,
we have the within-group contagion, with the infection rate
We have one more place where infection takes place, but this time it happens in the ether (or the mean-field)
On one hand,
GMEs provide a more accurate description of spreading processes when contagion depends not only on individual interactions but also on the internal dynamics of groups and how those groups are intertwined. When group sizes are heterogeneous and contagion is superlinear (i.e., $\nu > 1$), the system can exhibit mesoscopic localization: large groups disproportionately drive contagion, lowering the epidemic threshold. This can create hysteresis--where the long-term infection level depends on whether the outbreak began from a small or large seed. In real-world data (e.g., face-to-face contact networks), this implies that early momentum can lock in high infection levels, even if conditions later become unfavorable. Unlike annealed approximations, GMEs assume a quenched group topology--one that remains fixed as dynamics unfold--allowing the model to capture strong within-group correlations and persistent group influence. Groups here may represent contexts where local coordination shapes global dynamics, such as classrooms, teams, or households.
In this thesis, we build on those ideas by combining generalized master
equation models with evolutionary game theory, applied to group
dynamics. Here, instead of modeling contagion directly, we consider a
population of